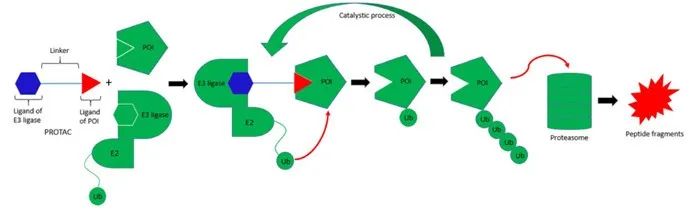
文章来源公众号:生物快评 作者:生物快评
慢病毒原理背景
HIV病毒颗粒和基因组
HIV病毒是属于envelope virus,这个envelope本意指的是信封上盖的蜡戳,用于封闭信件的作用,这里指的是病毒粒子外面有一层磷脂双分子脂质层。就是上图中的绿色部分,这样的有点是病毒的例子完全被双分子层的膜罩住了,那么暴露的抗原自然就少了。相对比non-envelope virus就需要病毒粒子直面宿主的免疫系统。
因为在水溶液中,人体的环境下,膜和膜融合不是天然发生的事情,需要克服膜周边水分子的排次作用,所以这样也给病毒和宿主细胞膜融合带来了另一个问题就是“感染入侵的问题”,这里需要进化出一套精密的“分子机器”,来完成病毒的膜和宿主细胞膜的融合。这些融合的完成需要依靠“配体-受体”作用来完成。
DOI:10.1016/j.bbamem.2016.02.007
病毒的蛋白质包括:
Gag: 这个gag是产生结构蛋白,gag这个名称怎么来的?一般在病毒学研究中,研究者对基因的命名充满了随意和简单直接,不像对人基因的命名要信达雅,这个鬼畜的缩写是group of antigen,意思是这个病毒蛋白主要激发人体产生抗体的,所以叫做“一组抗原”。
gag蛋白质产生MA/CA/NC。分别是MA: matrix蛋白,矩阵蛋白,结合包膜的作用,类似房屋的顶部支撑,。CA:capsid蛋白,包裹病毒粒子的作用,类似药物的胶囊壳。NC: nucleocapsid protein,NC是负责包裹病毒的核酸物质。最终通过
“俄罗斯套娃”一样的结构,得到封装好的HIV病毒粒子。
pol: pol蛋白是产生病毒的酶相关的蛋白:包括PR: 蛋白酶,RT:逆转录酶;IN: 整合酶(负责将DNA整合进入宿主基因组组)。
gp120和gp41,它们是一个复合体的两个部分,共同被称为包膜糖蛋白,是HIV病毒入侵人体细胞的“钥匙”,也是艾滋病药物和疫苗研究最重要的靶点之一。gp120:是暴露在外的“钩子”或“探头”,负责识别和初步结合目标细胞。gp41:是隐藏的“弹簧刀”或“锚”,负责膜融合,即刺穿细胞膜,让病毒核心进入细胞。
Rev:Rev is a transactivating protein that is essential to the regulation ofHIV-1 (and other lentiviral) protein expression。负责完整RNA从细胞核运输到细胞质中,完成病毒完整RNA和蛋白质的组装。
Tat: Trans-activator oftranscription(反式转录激活因子),大幅增强病毒基因的转录(将DNA模板转化为RNA的过程),没有Tat,HIV的复制效率会极低。
HIV的gp120/gp40包膜为VSV病毒的G蛋白
https://viralzone.expasy.org/5096
HIV病毒通过受体gp120结合受体CCR5、CD4之后,然后激动触发gp40暴露,gp40结构发生变化将gp40的疏水端插入到细胞膜上,介导病毒和细胞膜的融合。然后HIV capsid释放到了细胞质中,然后capsid经过激动蛋白结合并且在微管蛋白上运动,这样capsid运输到了核孔,最后向核孔内释放出病毒粒子中的酶和核酸。随后就发生了逆转录、整合等作用。
DOI 10.1074/jbc.M112.424432
HIV的gp160在病毒包装过程中经过宿主的蛋白酶水解得到gp120/gp41,两者是通过二硫键形成二聚体,然后在形成三聚体。Gp120糖基化程度高,且突变多,相当于亚稳态的gp41埋藏在gp120中,当gp120结合CD4CCR5等分子后,gp41改变而结合细胞膜,最后膜融合,病毒进入细胞。
Viruses 2024, 16(12), 1933; https://doi.org/10.3390/v16121933
再看VSV病毒通过G蛋白结合LDL-R,因为LDL-R天然的作用是结合脂蛋白,结合之后LDL-R和脂蛋白一起通过钙网蛋白被内吞,这样细胞就完成了脂质的摄入;VSV病毒的G蛋白就利用这个“运输的后门”,VSV的G蛋白结合LDL-R之后,细胞内吞进入endosome,VSV病毒进入到endosome中激发endosome中的质子泵而endosome提高了H+浓度,即pH下降。Endosome进一步转为lysosome,因为pH的下降,VSVG蛋白中一些组氨酸在pH中电荷改变导致VSVG结构变化,然后VSVG的疏水端暴露出来结合到lysosome的膜上,类似VSVG在lysosome中“撕开”膜,VSV病毒粒子释放到了细胞质中,随后VSV释放遗传物质,随后整个遗传物质复制、RNA的复制、转录,病毒组装均在细胞质中。
https://doi.org/10.1073/pnas.1618883114
目前慢病毒都将gp120/gp41复合物换成了VSV的病毒的包膜,那么这种改变其实除了改变HIV的靶向性,也改变了病毒的生命周期中的路径。
我们利用VSV.G包装的慢病毒通过逃逸endosome之后,慢病毒capsid依然比较完整,还可以接着依靠Dynein完成运输到细胞核。设想一下,如果VSV.G的融合逃逸能力和融合速度不足够,响应pH下降的速度较慢,那么慢病毒的capsid则会在lysosome中无法快速逃逸,最终LV壳就会破坏而进入不了细胞核。
第二代慢病毒和第三代慢病毒的区别
2nd vs 3rd Generation Lentiviral Systems
2nd vs 3rd Generation Lentiviral Systems | ||
Feature | 2nd Generation | 3rd Generation |
Transfer Plasmid | Can be packaged ONLY by a second generation packaging system that includes TAT | Can be packaged by both 2nd and 3rd generation packaging systems |
Packaging Plasmid | All on one plasmid: Gag, Pol, Rev, Tat | Two plasmids: one encoding Gag and Pol and another encoding Rev |
Envelope Plasmid | Interchangeable: usually encodes for VSV-G | Interchangeable: usually encodes for VSV-G |
Safety | Safe. Replication incompetent: Uses 3 separate plasmids encoding various HIV genes. | Safer. Replication incompetent and always SIN: Uses 4 plasmids instead of 3 and eliminates the requirement for Tat. |
LTR viral promoter | Wild type | Hybrid: 5'LTR is partially deleted and fused to a heterologous enhancer/promoter such as CMV or RSV |
通过上面的对比,我们可以发现,核心区别2nd慢病毒需要TAT,而3nd慢病毒不需要TAT;2nd中的gag、pol、rev、tat在一个质粒,而3nd慢病毒体系中,gagpol为同在一个质粒,而rev在一个质粒中。
那么这个Tat发挥了什么作用了呢?为什么3nd慢病毒就可以不依靠它了呢?
我们先来看看HIV 基因组是如何复制的?
https://viralzone.expasy.org/5061
这个图显示是HIV ssRNA,逆转录得到dsDNA的过程:
首选DNA->RNA转录,需要一个“引子”,比如真核生物转录所需要的引物酶(primase),病毒利用的是宿主的tRNA,tRNA结合到PBS上,tRNA充当了引物,从tRNA的5’-3’合成一段DNA,这段DNA和HIV的LTR的R-U5配对。
然后这个R-U5-tRNA的分子“跳跃”到HIV ssRNA的3’端继续依靠R配对,生成DNA,这里注意写到的RNA引物额外携带了新生的U5。新生的DNA和RNA形成DNA:RNA杂交链,随后RNA基本上被RNaseH降解,而在多聚嘧啶PPT剩余部分RNA充当引物,继续合成DNA,此时DNA携带了U3-R-U5,成为了完整的3‘LTR。随后这个DNA发生”跳跃”到HIV DNA的5‘端,通过形成PBS杂交DNA:DNA,继续得到dsDNA,此时dsDNA的两端都是完整的LTR。
然后我们要知道dsDNA如果整合到宿主基因组后,HIV的U5类似启动子来转录HIV DNA生成RNA。有两个问题:1)5‘LTR的U3启动HIV DNA 转录,但是3’ LTR的U3也会启动下游基因的转录,如果下游是抑癌基因则有风险。2)U3启动HIV DNA转录效率低下。
宿主依靠TAT提高转录水平:当病毒的DNA整合到宿主细胞的染色体后,细胞的转录机器(RNA聚合酶II)会开始读取病毒基因,合成病毒RNA。遇到“卡顿”:但在没有Tat的情况下,这个转录过程在病毒基因的起始阶段非常缓慢、低效,经常中途停止,只能产生非常短小的RNA片段。这就像一辆车刚启动就熄火,无法跑完全程。Tat蛋白会特异性结合到这些刚合成出来的、短小的病毒RNA的一个特殊区域,称为 TAR。Tat-TAR的结合就像一个“救援信号”,它会招募一大群宿主细胞的辅助因子(如P-TEFb)。这个复合体(Tat + TAR + 宿主因子)能极大地提升RNA聚合酶的“工作效率”。
举例子:
HIV野生型的基因组ssRNA结构
这里需要特别注意R-U5,5端独特序列(Unique to the 5’ end);U3-R中的3‘ 端独特序列(Unique to the 3’ end)。
当HIV基因组整合到宿主基因组后,前病毒就变成了宿主细胞的一部分(即DNA序列)。这时,dsDNA序列的5端是U3-R-U5-genome-U3-R-U5(见上HIV 基因组逆转录的示意图).
HIV整合后转录时,U3区域发挥转录激活启动子的作用:U3区域内包含了许多关键的调控序列,可以结合宿主细胞的转录因子(如NF-κB, SP1等),并且这个转录活性依然不足够,病毒自身的Tat蛋白(通过TAR元件,位于R区)。当这些因子结合到U3区域后,它们会招募细胞的RNA聚合酶II,启动病毒基因的转录。
第二代慢病毒的质粒图谱
Addgene的pWPT-GFP (Plasmid #12255)
这个是二代2nd病毒目的载体。5’LTR是完整的LTR(即U3-R-U5),truncated 3’LTR(R-U5)缺少U3。如果这个载体进行慢病毒包装,那么就相当于HIV在宿主基因中转录出来ssRNA包装进入慢病毒中,ssRNA的结构是:R-U5-GOI(Gene of Interest)-R。在这个过程中,需要完全依靠包装载体自带的5’LTR的U3启动转录,所需需要加入TAT协助转录得到更多的ssRNA,提高病毒产量。
当这个ssRNA经过逆转录后得到dsRNA,其结构是:R-U5-GOI-R-U5。这样目的基因就依靠自身的启动子转录(比如这里的EF1A),而3’LTR并没有U3,所以也不会对下游基因产生激活转录。
第三代慢病毒的质粒图谱
Addgene的pLVpuro-CMV-N-EGFP(Plasmid #122848)
这个是二代3nd病毒目的载体。5’LTR是truncated LTR(即R-U5),3’LTR(U3.truncated-R-U5),根据生物过程,这个质粒转染进入细胞,因为5LTR缺少U3,他是无法转录出ssRNA的,那么怎么办呢?这里额外加入外源强启动子RSV-promoter等转录出的ssRNA。这个ssRNA的结构是R3-U5-GOI-U3.truncated-R,因为这个U3.truncated很短已经缺少激活能力,所有也不会激活下游基因。因为慢病毒的5’LTR已经缺少U3且用了外源强启动子, 那么自然也就不需要TAT了。